
Authors
Larisa Rudenko 1,†, Irina Kiseleva 1,†, Elena Krutikova 1, Ekaterina Stepanova 1,*, Irina Isakova-Sivak 1, Svetlana Donina 1, Andrey Rekstin 1, Maria Pisareva 1, Ekaterina Bazhenova 1, Tatiana Kotomina 1, Anastasia Katelnikova 2, Arman Muzhikyan 2, Valery Makarov 2, Erin Grace Sparrow 3 and Guido Torelli 3
1 Institute of Experimental Medicine, St. Petersburg 197376, Russia
2 Institute of Preclinical Research Ltd., St. Petersburg 188663, Russia
3 World Health Organization, 1211 Geneva, Switzerland
* Correspondence: fedorova.iem@gmail.com; Tel.: +7-812-234-4292
† These authors contributed equally to this work as first authors.
Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Ethics Statement
2.3. Animals
2.4. Study Design
2.5. Clinical Signs and Morbidity Outcomes
2.6. Determination of Virus Load in Embryonated Chicken Eggs
2.7. PCR-Based H7N9 Vaccine Virus Detection
2.8. Hemagglutination Inhibition Assay
2.9. Determination of Influenza-Specific Ferret IgG/IgA
2.10. Necropsy
2.11. Histopathology
2.12. Testing of Human Serum Samples
2.13. Statistics
3. Results
3.1. Clinical Observations in Ferrets
3.1.1. Body Weight
3.1.2. Clinical Signs
3.1.3. Body Temperature
3.2. Vaccine Virus Replication
Notably, no live virus was found in nasal washes of ferrets in groups 1 and 2 after the second dose of H7N9 LAIV. None of the animals in groups 1–3 had live vaccine virus in the upper airways (Table 2). These data indicate that the single dose of H7N9 vaccine prevented vaccine virus replication after revaccination.
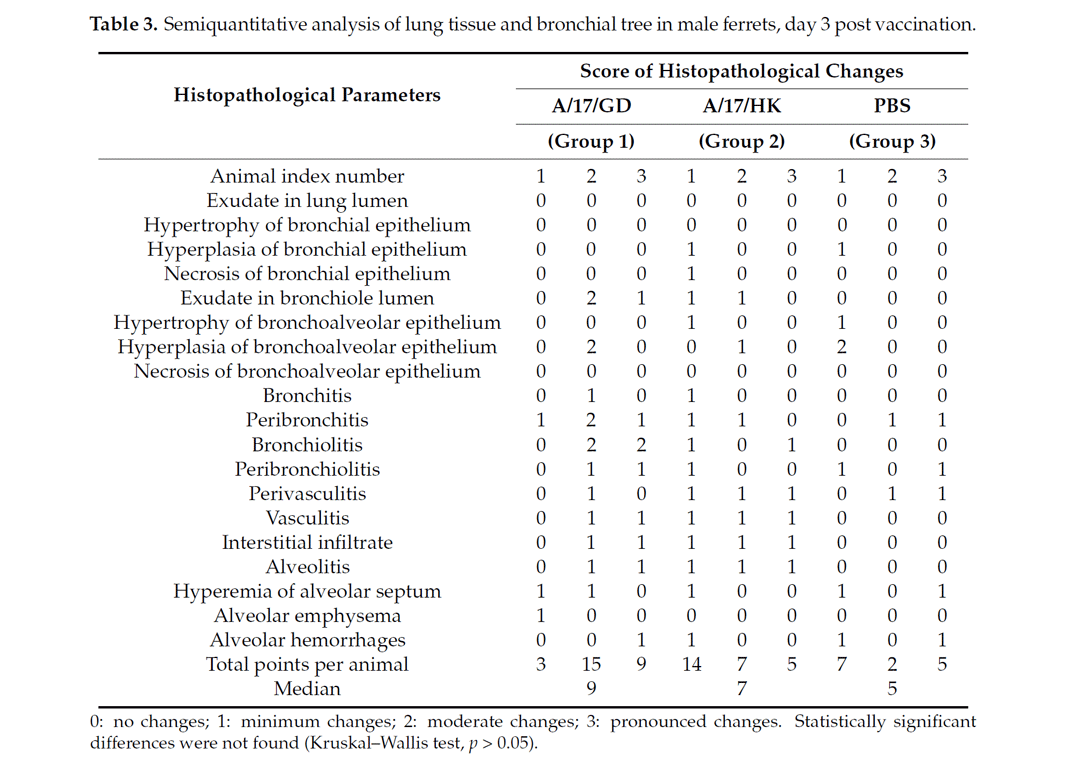
3.3. Pathomorphological Examination of Trachea and Lungs
3.4. Antibody Response to H7N9 LAIV
3.5. Heterologous Antibody Immune Response of Healthy Volunteers to H7N9 Viruses
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Disclaimer
Conflicts of Interest
References
- Kluska, V.; Macku, M.; Mensik, J. Demonstration of antibodies against swine influenza viruses in man. Cesk. Pediatr. 1961, 16, 408–414. [Google Scholar] [PubMed]
- Widdowson, M.-A.; Bresee, J.S.; Jernigan, D.B. The global threat of animal influenza viruses of zoonotic concern: Then and now. J. Infect. Dis. 2017, 216, S493–S498. [Google Scholar] [CrossRef] [PubMed]
- Joseph, U.; Su, Y.C.F.; Vijaykrishna, D.; Smith, G.J.D. The ecology and adaptive evolution of influenza A interspecies transmission. Influenza Other Respir. Viruses 2017, 11, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Paules, C.; Subbarao, K. Influenza. Lancet 2017, 390, 697–708. [Google Scholar] [CrossRef]
- World Health Organization. Human Infection with Influenza A(H7N9) Virus in China. Available online: http://www.who.int/csr/don/2013_04_01/en/(accessed on 25 August 2018).
- World Health Organization. Antigenic and Genetic Characteristics of Zoonotic Influenza Viruses and Development of Candidate Vaccine Viruses for Pandemic Preparedness. February 2018. Available online: http://www.who.int/influenza/vaccines/virus/201802_zoonotic_vaccinevirusupdate.pdf?ua=1 (accessed on 25 August 2018).
- World Health Organization. WHO Recommendations on the Composition of Influenza Virus Vaccines. Available online: http://www.who.int/influenza/vaccines/virus/recommendations/en/ (accessed on 25 August 2018).
- World Health Organization. Antigenic and Genetic Characteristics of Zoonotic Influenza Viruses and Development of Candidate Vaccine Viruses for Pandemic Preparedness. March 2017. Available online: http://www.who.int/influenza/vaccines/virus/201703_zoonotic_vaccinevirusupdate.pdf?ua=1(accessed on 25 August 2018).
- Centers for Disease Control and Prevention. Summary of Influenza Risk Assessment Tool (IRAT) Results. Available online: https://www.cdc.gov/flu/pandemic-resources/monitoring/irat-virus-summaries.htm (accessed on 25 August 2018).
- Penttinen, P.M.; Friede, M.H. Decreased effectiveness of the influenza A(H1N1) pdm09 strain in live attenuated influenza vaccines: An observational bias or a technical challenge? Eurosurveillance 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Cox, N.J.; Hickling, J.; Jones, R.; Rimmelzwaan, G.F.; Lambert, L.C.; Boslego, J.; Rudenko, L.; Yeolekar, L.; Robertson, J.S.; Hombach, J.; et al. Report on the second WHO integrated meeting on development and clinical trials of influenza vaccines that induce broadly protective and long-lasting immune responses: Geneva, Switzerland, 5–7 May 2014. Vaccine 2015, 33, 6503–6510. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, L.; Yeolekar, L.; Kiseleva, I.; Isakova-Sivak, I. Development and approval of live attenuated influenza vaccines based on Russian master donor viruses: Process challenges and success stories. Vaccine 2016, 34, 5436–5441. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, L.; Desheva, J.; Korovkin, S.; Mironov, A.; Rekstin, A.; Grigorieva, E.; Donina, S.; Gambaryan, A.; Katlinsky, A. Safety and immunogenicity of live attenuated influenza reassortant H5 vaccine (phase I–II clinical trials). Influenza Other Respir. Viruses 2008, 2, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, L.; Isakova-Sivak, I.; Naykhin, A.; Kiseleva, I.; Stukova, M.; Erofeeva, M.; Korenkov, D.; Matyushenko, V.; Sparrow, E.; Kieny, M.-P. H7N9 live attenuated influenza vaccine in healthy adults: a randomised, double-blind, placebo-controlled, phase 1 trial. Lancet Infect. Dis. 2016, 16, 303–310. [Google Scholar] [CrossRef]
- Rudenko, L.; Kiseleva, I.; Naykhin, A.N.; Erofeeva, M.; Stukova, M.; Donina, S.; Petukhova, G.; Pisareva, M.; Krivitskaya, V.; Grudinin, M.; et al. Assessment of human immune responses to H7 avian influenza virus of pandemic potential: Results from a placebo–controlled, randomized double–blind phase I study of live attenuated H7N3 influenza vaccine. PLoS ONE2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, L.; van den Bosch, H.; Kiseleva, I.; Mironov, A.; Naikhin, A.; Larionova, N.; Bushmenkov, D. Live attenuated pandemic influenza vaccine: Clinical studies on A/17/California/2009/38 (H1N1) and licensing of the Russian-developed technology to WHO for pandemic influenza preparedness in developing countries. Vaccine 2011, 29. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, L.; Kiseleva, I.; Stukova, M.; Erofeeva, M.; Naykhin, A.; Donina, S.; Larionova, N.; Pisareva, M.; Krivitskaya, V.; Flores, J. Russian LAIV trial study group clinical testing of pre-pandemic live attenuated A/H5N2 influenza candidate vaccine in adult volunteers: Results from a placebo-controlled, randomized double-blind phase I study. Vaccine 2015, 33, 5110–5117. [Google Scholar] [CrossRef] [PubMed]
- Isakova-Sivak, I.; Stukova, M.; Erofeeva, M.; Naykhin, A.; Donina, S.; Petukhova, G.; Kuznetsova, V.; Kiseleva, I.; Smolonogina, T.; Dubrovina, I.; et al. H2N2 live attenuated influenza vaccine is safe and immunogenic for healthy adult volunteers. Hum. Vaccines Immunother. 2015, 11, 970–982. [Google Scholar] [CrossRef] [PubMed]
- Wareing, M.D.; Marsh, G.A.; Tannock, G.A. Preparation and characterisation of attenuated cold-adapted influenza A reassortants derived from the A/Leningrad/134/17/57 donor strain. Vaccine 2002, 20, 2082–2090. [Google Scholar] [CrossRef]
- Shcherbik, S.; Pearce, N.; Kiseleva, I.; Larionova, N.; Rudenko, L.; Xu, X.; Wentworth, D.E.; Bousse, T. Implementation of new approaches for generating conventional reassortants for live attenuated influenza vaccine based on Russian master donor viruses. J. Virol. Methods 2016, 227, 33–39. [Google Scholar] [CrossRef] [PubMed]
- De Jonge, J.; Isakova-Sivak, I.; van Dijken, H.; Spijkers, S.; Mouthaan, J.; de Jong, R.; Smolonogina, T.; Roholl, P.; Rudenko, L. H7N9 live attenuated influenza vaccine is highly immunogenic, prevents virus replication, and protects against severe bronchopneumonia in ferrets. Mol. Ther. 2016, 24, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Global Pandemic Action Plan to Increase Supply of Pandemic Influenza Vaccine. Available online: http://www.who.int/csr/resources/publications/influenza/CDS_EPR_GIP_2006_1.pdf (accessed on 25 August 2018).
- Isakova-Sivak, I.; Chen, L.-M.; Matsuoka, Y.; Voeten, J.T.M.; Kiseleva, I.; Heldens, J.G.M.; van den Bosch, H.; Klimov, A.; Rudenko, L.; Cox, N.J.; et al. Genetic bases of the temperature-sensitive phenotype of a master donor virus used in live attenuated influenza vaccines: A/Leningrad/134/17/57 (H2N2). Virology 2011, 412, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes. Official Journal of the European Union. L 276/33. Available online: http://data.europa.eu/eli/dir/2010/63/oj/eng (accessed on 25 August 2018).
- World Health Organization. WHO Manual on Animal Influenza Diagnosis and Surveillance. Available online: http://www.wpro.who.int/emerging_diseases/documents/docs/manualonanimalaidiagnosisandsurveillance.pdf(accessed on 25 August 2018).
- Zang, Y.; Du, D.; Ge, P.; Xu, Y.; Liu, X.; Zhang, Y.; Su, W.; Kiseleva, I.; Rudenko, L.; Xu, F.; et al. Development of one-step real-time PCR assay for titrating trivalent live attenuated influenza vaccines. Hum. Vaccines Immunother. 2014, 10, 3642–3648. [Google Scholar] [CrossRef] [PubMed]
- Suguitan, A.L.; Cheng, X.; Wang, W.; Wang, S.; Jin, H.; Lu, S. Influenza H5 hemagglutinin DNA primes the antibody response elicited by the live attenuated influenza A/Vietnam/1203/2004 vaccine in ferrets. PLoS ONE2011, 6, e21942. [Google Scholar] [CrossRef] [PubMed]
- Widjojoatmodjo, M.N.; Boes, J.; van Bers, M.; van Remmerden, Y.; Roholl, P.J.; Luytjes, W. A highly attenuated recombinant human respiratory syncytial virus lacking the G protein induces long-lasting protection in cotton rats. Virol. J. 2010, 7, 114. [Google Scholar] [CrossRef] [PubMed]
- Gaydos, J.C.; Top, F.H.; Hodder, R.A.; Russell, P.K. Swine influenza A outbreak, Fort Dix, New Jersey, 1976. Emerg. Infect. Dis. 2006, 12, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Creanga, A.; Hang, N.L.K.; Cuong, V.D.; Nguyen, H.T.; Phuong, H.V.M.; Thanh, L.T.; Thach, N.C.; Hien, P.T.; Tung, N.; Jang, Y.; et al. Highly pathogenic avian influenza A(H5N1) viruses at the animal-human interface in Vietnam, 2003–2010. J. Infect. Dis. 2017, 216, S529–S538. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Feng, Z.; Shu, Y.; Yu, H.; Zhou, L.; Zu, R.; Huai, Y.; Dong, J.; Bao, C.; Wen, L.; et al. Probable limited person-to-person transmission of highly pathogenic avian influenza A (H5N1) virus in China. Lancet Lond. Engl. 2008, 371, 1427–1434. [Google Scholar] [CrossRef]
- World Health Organization. Assessment of Risk Associated with Influenza A(H5N8) Virus. Available online: http://www.who.int/influenza/human_animal_interface/avian_influenza/riskassessment_AH5N8_201611/en/ (accessed on 25 August 2018).
- Zhang, R.; Chen, T.; Ou, X.; Liu, R.; Yang, Y.; Ye, W.; Chen, J.; Yao, D.; Sun, B.; Zhang, X.; et al. Clinical, epidemiological and virological characteristics of the first detected human case of avian influenza A(H5N6) virus. Infect. Genet. Evol. 2016, 40, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.; Gao, R.; Lv, Q.; Huang, S.; Zhou, Z.; Yang, L.; Li, X.; Zhao, X.; Zou, X.; Tong, W.; et al. Human infection with a novel, highly pathogenic avian influenza A (H5N6) virus: Virological and clinical findings. J. Infect.2016, 72, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.-H.; Yang, J.-R.; Wu, H.-S.; Chang, M.-C.; Lin, J.-S.; Lin, C.-Y.; Liu, Y.-L.; Lo, Y.-C.; Yang, C.-H.; Chuang, J.-H.; et al. Human infection with avian influenza A H6N1 virus: An epidemiological analysis. Lancet Respir. Med. 2013, 1, 771–778. [Google Scholar] [CrossRef]
- Tweed, S.A.; Skowronski, D.M.; David, S.T.; Larder, A.; Petric, M.; Lees, W.; Li, Y.; Katz, J.; Krajden, M.; Tellier, R.; et al. Human illness from avian influenza H7N3, British Columbia. Emerg. Infect. Dis. 2004, 10, 2196–2199. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Martinez, I.; Balish, A.; Barrera-Badillo, G.; Jones, J.; Nuñez-García, T.E.; Jang, Y.; Aparicio-Antonio, R.; Azziz-Baumgartner, E.; Belser, J.A.; Ramirez-Gonzalez, J.E.; et al. Highly pathogenic avian influenza A(H7N3) virus in poultry workers, Mexico, 2012. Emerg. Infect. Dis. 2013, 19, 1531–1534. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Bridges, C.B.; Katz, J.M.; Tumpey, T.M. Past, present, and possible future human infection with influenza virus A subtype H7. Emerg. Infect. Dis. 2009, 15, 859–865. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Influenza at the Human–Animal Interface. Summary and Assessment. Available online: http://www.who.int/influenza/human_animal_interface/Influenza_Summary_IRA_HA_interface_02_03_2018.pdf (accessed on 26 August 2018).
- Fouchier, R.A.M.; Schneeberger, P.M.; Rozendaal, F.W.; Broekman, J.M.; Kemink, S.A.G.; Munster, V.; Kuiken, T.; Rimmelzwaan, G.F.; Schutten, M.; Van Doornum, G.J.J.; et al. Avian influenza A virus (H7N7) associated with human conjunctivitis and a fatal case of acute respiratory distress syndrome. Proc. Natl. Acad. Sci. USA 2004, 101, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, M.; Wilbrink, B.; Conyn, M.; Natrop, G.; van der Nat, H.; Vennema, H.; Meijer, A.; van Steenbergen, J.; Fouchier, R.; Osterhaus, A.; et al. Transmission of H7N7 avian influenza A virus to human beings during a large outbreak in commercial poultry farms in the Netherlands. Lancet Lond. Engl. 2004, 363, 587–593. [Google Scholar] [CrossRef]
- Belser, J.A.; Creager, H.M.; Zeng, H.; Maines, T.R.; Tumpey, T.M. Pathogenesis, transmissibility, and tropism of a highly pathogenic avian influenza A(H7N7) virus associated with human conjunctivitis in Italy, 2013. J. Infect. Dis. 2017, 216, S508–S511. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ren, X.; Li, L.; Xiao, Y.; Dong, J.; Sun, L.; Zhu, Y.; Yang, F.; Zhang, X.; Jin, Q. Rapid genome sequencing and characterization of novel avian-origin influenza A H7N9 virus directly from clinical sample by semiconductor sequencing. J. Clin. Virol. 2015, 73, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Cao, B.; Hu, Y.; Feng, Z.; Wang, D.; Hu, W.; Chen, J.; Jie, Z.; Qiu, H.; Xu, K.; et al. Human infection with a novel avian-origin influenza A (H7N9) virus. N. Engl. J. Med. 2013, 368, 1888–1897. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liang, W.; Yang, S.; Wu, N.; Gao, H.; Sheng, J.; Yao, H.; Wo, J.; Fang, Q.; Cui, D.; et al. Human infections with the emerging avian influenza A H7N9 virus from wet market poultry: Clinical analysis and characterisation of viral genome. Lancet 2013, 381, 1916–1925. [Google Scholar] [CrossRef]
- Ding, H.; Xie, L.; Sun, Z.; Kao, Q.-J.; Huang, R.-J.; Yang, X.-H.; Huang, C.; Wen, Y.-Y.; Pan, J.-C.; Pu, X.-Y.; et al. Epidemiologic characterization of 30 confirmed cases of human infection with avian influenza A(H7N9) virus in Hangzhou, China. BMC Infect. Dis. 2014, 14, 175. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Lim, W.; Suzuki, T.; Suzuki, Y.; Kida, H.; Nishimura, S.I.; Tashiro, M. Characterization of a human H9N2 influenza virus isolated in Hong Kong. Vaccine 2001, 20, 125–133. [Google Scholar] [CrossRef]
- Peiris, M.; Yuen, K.Y.; Leung, C.W.; Chan, K.H.; Ip, P.L.; Lai, R.W.; Orr, W.K.; Shortridge, K.F. Human infection with influenza H9N2. Lancet 1999, 354, 916–917. [Google Scholar] [CrossRef]
- Liu, M.; Li, X.; Yuan, H.; Zhou, J.; Wu, J.; Bo, H.; Xia, W.; Xiong, Y.; Yang, L.; Gao, R.; et al. Genetic diversity of avian influenza A (H10N8) virus in live poultry markets and its association with human infections in China. Sci. Rep. 2015, 5, 7632. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yuan, H.; Gao, R.; Zhang, J.; Wang, D.; Xiong, Y.; Fan, G.; Yang, F.; Li, X.; Zhou, J.; et al. Clinical and epidemiological characteristics of a fatal case of avian influenza A H10N8 virus infection: A descriptive study. Lancet 2014, 383, 714–721. [Google Scholar] [CrossRef]
- World Health Organization. WHO Global Influenza Preparedness Plan WHO/CDS/CSR/GIP/2005.5. Available online: http://www.who.int/csr/resources/publications/influenza/en/WHO_CDS_CSR_GIP_2005_5.pdf (accessed on 25 August 2018).
- World Health Organization. Tables on clinical evaluation of influenza vaccines. Pandemic and potentially pandemic influenza vaccines. Available online: http://www.who.int/entity/immunization/diseases/influenza/Table_clinical_evaluation_influenza_pandemic.xlsx (accessed on 25 August 2018).
- He, X.-S.; Holmes, T.H.; Zhang, C.; Mahmood, K.; Kemble, G.W.; Lewis, D.B.; Dekker, C.L.; Greenberg, H.B.; Arvin, A.M. Cellular immune responses in children and adults receiving inactivated or live attenuated influenza vaccines. J. Virol. 2006, 80, 11756–11766. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, L.G.; Slepushkin, A.N.; Monto, A.S.; Kendal, A.P.; Grigorieva, E.P.; Burtseva, E.P.; Rekstin, A.R.; Beljaev, A.L.; Bragina, V.E.; Cox, N. Efficacy of live attenuated and inactivated influenza vaccines in schoolchildren and their unvaccinated contacts in Novgorod, Russia. J. Infect. Dis. 1993, 168, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Barría, M.I.; Garrido, J.L.; Stein, C.; Scher, E.; Ge, Y.; Engel, S.M.; Kraus, T.A.; Banach, D.; Moran, T.M. Localized mucosal response to intranasal live attenuated influenza vaccine in adults. J. Infect. Dis. 2013, 207, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Tamura, S.; Tanimoto, T.; Kurata, T. Mechanisms of broad cross-protection provided by influenza virus infection and their application to vaccines. Jpn. J. Infect. Dis. 2005, 58, 195–207. [Google Scholar] [PubMed]
- Hoft, D.F.; Babusis, E.; Worku, S.; Spencer, C.T.; Lottenbach, K.; Truscott, S.M.; Abate, G.; Sakala, I.G.; Edwards, K.M.; Creech, C.B.; et al. Live and inactivated influenza vaccines induce similar humoral responses, but only live vaccines induce diverse T-cell responses in young children. J. Infect. Dis. 2011, 204, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Ilyushina, N.A.; Haynes, B.C.; Hoen, A.G.; Khalenkov, A.M.; Housman, M.L.; Brown, E.P.; Ackerman, M.E.; Treanor, J.J.; Luke, C.J.; Subbarao, K.; et al. Live attenuated and inactivated influenza vaccines in children. J. Infect. Dis. 2015, 211, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.F.; Hoen, A.G.; Ilyushina, N.A.; Brown, E.P.; Ackerman, M.E.; Wieland-Alter, W.; Connor, R.I.; Jegaskanda, S.; Rosenberg-Hasson, Y.; Haynes, B.C.; et al. Correlates of immunity to influenza as determined by challenge of children with live, attenuated influenza vaccine. Open Forum Infect. Dis. 2016, 3. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Zhang, Q.; Gu, C.; Shi, J.; Deng, G.; Ma, S.; Liu, J.; Chen, P.; Guan, Y.; Jiang, Y.; et al. A live attenuated vaccine prevents replication and transmission of H7N9 virus in mammals. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]